 |
| Manos dibujándose de MC Escher (origen) |
En la entrada "
De la forma" comentamos la importancia evolutiva que tiene la forma de las bacterias y que la morfología bacteriana era mucho más diversa de lo que en un principio se pensaba.
Pero, ¿cómo se forma una bacteria? La verdad es que sabemos más bien poco ya que lo que se conoce se basa en el estudio de unas cuantas bacterias: los cocos
Streptococcus pneumoniae y
Staphylococcus aureus; los bacilos
Escherichia coli y
Bacillus subtilis; las curvas y espirales formadas por
Helicobacter pylori,
Caulobacter crecentus o
Borrelia burgdorferi. Del resto de morfologías no se conoce mucho. Así que quizás habría que replantearse la pregunta y elaborar alguna más simple que sí podamos contestar. Por ejemplo:
En el aspecto de la morfogénesis ¿qué tienen en común todas las bacterias?
- Todas tienen paredes de peptidoglicano (salvo los micoplasmas y los termoplasmas) y la forma viene dada por cómo está construida dicha macromolécula a nivel molecular. Así que no es de extrañar que el equipamiento bioquímico responsable de la síntesis de esa pared se haya conservado entre los diversos phyla bacterianos. A nivel molecular, ese equipamiento enzimático se coloca en una determinada “zona” de la bacteria y es allí donde ejerce su función. Es lo que podría denominarse una "síntesis zonal"
- La maquinaria de síntesis de la pared debe de asegurar que se mantenga la forma y que se incremente el tamaño de la bacteria gracias a la elongación, pero también debe permitir la reproducción celular mediante la septación. Basta cambiar mínimamente la orientación de la síntesis de peptidoglicano para pasar de elongar a septar. Lo que sí es necesario es que ambos procesos deben de estar coordinados tanto en el espacio como en el tiempo para asegurar la supervivencia de la bacteria.
 |
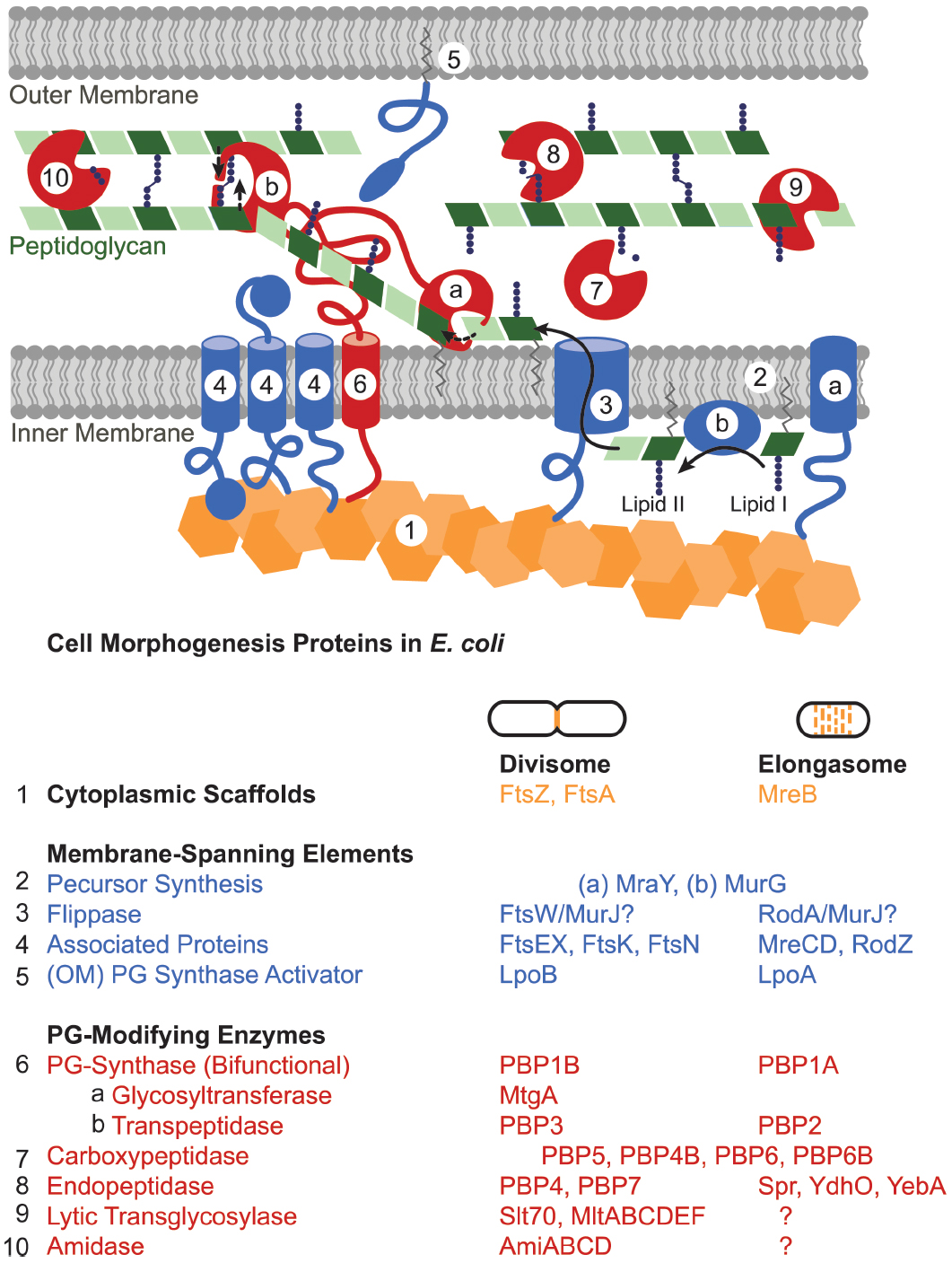
| Maquinaria de síntesis de peptidoglicano en la bacteria Escherichia coli. La elongación la lleva a cabo el elongasoma y la división el divisioma. En ambos casos se necesita un armazón citoplasmático que va a dirigir la localización de la síntesis del peptidoglicano (color naranja) y que reclutaran los diversos componentes como los elementos de membrana que incluyen proteínas reguladoras y sintetizadoras de elementos precursores (color azul) y proteínas sintetizadoras y modificadoras del peptidoglicano (color rojo). De todas estas proteínas, las más conservadas en todos los grupos bacterianos son FtsZ (naranja) y FtsK (azul-4). Origen de la imagen: Amelia M. Randich e Yves V. Brun 2015 |
Podríamos decir que esas son las "reglas básicas" que toda bacteria tiene que seguir. Quizás, cada grupo bacteriano tenga además algunas reglas específicas que expliquen las diferencias morfológicas. O quizás, uno podría esperar que si dos bacterias tienen una forma bacilar será porque ambas siguen el mismo juego de reglas. El mejor ejemplo es
Escherichia coli (Gram negativa) y
Bacillus subtilis (Gram positiva). Ambas realizan la elongación gracias a que sus equipos enzimáticos se colocan en unas zonas que siguen una pauta espiral y así sintetizan el nuevo peptidoglicano siguiendo dicha pauta.
 |
| Incorporación del peptidoglicano en diferentes bacterias morfológicamente distintas. En A se representa la elongación, en B la división. Tanto en Escherichia coli como en Bacillus subtilis, las zonas de elongación se disponen a lo largo de una espiral. Cuando las células se van a dividir la síntesis zonal se localiza solamente en el centro de la bacteria. En el caso de Streptococcus pneumoniae, la elongación se produce al insertar nuevo material en el llamado anillo ecuatorial. Según se va insertando el nuevo material los anillos resultantes se separana señalando el futuro lugar de septación. En el caso de Staphylococcus aureus no parece haber elongación, tan sólo síntesis de peptidoglicano septal. Origen de la imagen Dirk-Jan Scheffers1 y Mariana G. Pinho. 2005. |
El dato de que dos bacterias tan alejadas filogenéticamente entre sí, han resuelto el mismo problema de la misma manera puede apuntar a que dicha "solución" apareció hace mucho tiempo. Pero cuando se observa con detalle lo que ocurre en otros grupos taxonómicos (ver el
árbol filogenético de la 1ª parte), lo que a primera vista parece una
conservación evolutiva es más bien una
convergencia evolutiva. Entre una y otra especie nos encontramos con una miríada de morfologías. Observar una convergencia evolutiva nos indica otra cosa distinta: si se reposiciona la maquinaria de síntesis zonal se pueden generar un gran diversidad de formas bacterianas todas ellas derivadas de la morfología bacilar. Por ejemplo, si la síntesis zonal se restringe a uno sólo de los polos podemos obtener una prosteca como la de
Caulobacter crescentus. En cambio, si la síntesis zonal se distribuye a lo largo de la longitud de un filamento se pueden conseguir las ramificaciones que se observan en las Actinobacterias. Bastaría con usar las reglas básicas de diferente forma para obtener las diversas morfologías bacterianas.
 |
| Síntesis zonal del peptidoglicano y morfología bacteriana. En las Alfaproteobacterias prostecadas basta reposicionar la maquinaria de síntesis del peptidoglicano (en verde) para que aparezcan diferentes fenotipos. A: Caulobacter crescentus prosteca polar. B: Asticcacaulis excentricus, prosteca subpolar. C: Asticcacaulis biprosthecum prostecas bipolares. En D tenemos el fenotipo ramificado de la actinobacteria Streptomyces coelicolor. Origen de la imagen: David T. Kysela et al. 2016. |
Combinando tecnologías genómicas de una sola célula (single-cell genomics) con las tecnologías de microscopía de alta resolución, el grupo de Yves Brunn ha conseguido estudiar la evolución de la prosteca entre las especies pertenecientes a los Caulobacterales. Observando el árbol filogenético que se muestra más abajo, puede verse que primero apareció la prosteca polar (presente en
Caulobacter crescentus). En algunas especies esa prosteca se ha convertido en una zona especializada en la reproducción por gemación (
Hirschia baltica) pero en otras lo que ha pasado es que se ha perdido y la bacteria ha vuelto a una morfología bacilar (
C. segnis). En otras especies, la síntesis zonal se reposicionó de manera supolar (
Asticcacaulis excentricus) y a partir de este grupo volvió a reposicionarse a bilateral aumentando su número (
A. biprosthecum). Al examinar con más detalle dicha transición, lo que se encontraron es que hay una proteína, denominada SpmX, que es la responsable de coordinar la síntesis del peptidoglicano en la prosteca. Dicha proteína tiene una región que es diferente en cada una de esas especies y que determina la localización de la síntesis zonal
 |
| Arbol filogenético de los Caulobacterales y morfología celular. 1. prosteca bilateral, Asticcacaulis biprosthecum (Chao Jiang, Stanford University). 2. prosteca subpolar, Asticcacaulis excentricus (Chao Jiang, Stanford University). 3. prosteca polar, Caulobacter crescentus (Paul Caccamo, Indiana University). 4. prosteca polar, Maricaulis maris (Patrick Viollier, University of Geneva). 5. prosteca polar pequeña, Brevundimonas subvibriodes (Brynn Heckel, Indiana University). 6. Prosteca polar para reproducción por gemación, Hirschia baltica. Origen de la imagen: David T. Kysela et al. 2016. |
¿Podría explicarse la gran diversidad de morfologías bacterianas en base a la síntesis zonal de peptidoglicano? Pues es muy probable que la combinación de métodos genómicos con las nuevas tecnologías microscópicas den lugar a nuevas aproximaciones experimentales que permitan dar respuesta a esa pregunta y estudiar porqué las bacteria son como son e incluso abordar el problema de relacionar la morfogénesis bacteriana y las fuerzas de selección que actúan sobre ella. Puede afirmarse que este campo de investigación va tomando forma
Esta entrada y su primera parte han sido traducidas y publicadas en el blog
Small Things Considered

{kind=link}


No hay comentarios:
Publicar un comentario