Si se realizara una encuesta a todos aquellos que estudian algún aspecto de la Biología sobre cual es la disciplina menos atractiva, es muy probable que la Taxonomía ocupe uno de los primeros puestos. Se supone que es un conjunto de reglas para nombrar a los seres vivos, pero dichas reglas no parecen muy intuitivas y para colmo cambian cada cierto tiempo y al final parece que se discute sobre el sexo de los ángeles en lugar de sobre las formas vivas.
Sin embargo, la Taxonomía es el oficio más antiguo del mundo o al menos eso podemos leer en la Biblia, porque tras la creación de los animales se lee:
Génesis 2.20: El hombre puso un nombre a todos los animales domésticos, a todas las aves del cielo y a todos los animales del campo; pero entre ellos no encontró la ayuda adecuada.
Después de la creación de la mujer surgió la anatomía comparada y comenzaron las primeras discusiones taxonómicas, una maldición que aún perdura.

Sin embargo, la Taxonomía es el oficio más antiguo del mundo o al menos eso podemos leer en la Biblia, porque tras la creación de los animales se lee:
Génesis 2.20: El hombre puso un nombre a todos los animales domésticos, a todas las aves del cielo y a todos los animales del campo; pero entre ellos no encontró la ayuda adecuada.
Después de la creación de la mujer surgió la anatomía comparada y comenzaron las primeras discusiones taxonómicas, una maldición que aún perdura.

Entrega de certificados de asistencia en el Primer Congreso Mundial de Taxonomía
De todas formas hay que comprender a los taxónomos. Las reglas taxonómicas son el fruto de un convenio entre los científicos, y si hay algo frágil en este mundo es precisamente un convenio científico. Pero ahí no acaba la cosa. La taxonomía trata de clasificar a las especies biológicas. Y esa es la madre del cordero, porque ¿qué es una especie biológica?
Si uno trabaja con animales como hizo Adán la cosa parece sencilla. Un león es un león, un perro es un perro y un ratón es un ratón. Pero la inteligente Eva le hizo notar algo bastante peculiar. Las especies "león", "perro" y "ratón" necesitan de dos individuos, un macho y una hembra, para reproducirse y generar más leones, perros y ratones. Y eso no sólo pasaba con los animales, también con las plantas. Llegamos así a la definición más conocida de especie biológica: grupo de organismos capaces de entrecruzarse y de producir descendencia fértil.
Eso significa que hay que cambiar o ampliar la definición de "especie biológica", lo que quiere decir que hay que montar un congreso de taxonomía, poner un montón de señores a discutir y después de unos días publicar un acta del congreso donde se describirá el nuevo convenio científico sobre lo que es una especie biológica. Según el libro "Brock. Biología de los microorganismos", una especie microbiológica es una colección de cepas que comparten un gran número de propiedades importantes pero difieren en una o más propiedades significativas de otras colecciones de cepas. Siendo sinceros a mi no me parece ni corta ni sencilla, pero es lo que hay.
Y ahora viene la parte más divertida. Podría pensarse que un taxónomo es como un bibliotecario o como el encargado de un vídeo-club. El taxónomo clasifica seres vivos como el librero clasifica libros o el encargado del vídeo-club, películas. Pero hay una diferencia fundamental entre el primero y los otros dos. Los seres vivos evolucionan, cambian en el tiempo.
Uno puede pensar que lo mismo ocurre con los libros y las películas. Antes las películas se alquilaban en formato de video y ahora sólo hay DVD, dentro de poco serán en Blu-ray. Pero lo que ha cambiado es el soporte, no la información. En los seres vivos, la evolución significa cambio en la información. Y si cambia lo información significa que esa "especie" se ha transformado en "otra especie". Para hacernos una idea, el cambio evolutivo biológico es como si en una biblioteca tenemos un ejemplar de "El Quijote" y tras pasar un tiempo ese libro se ha transformado y ha pasado de ser una novela a un tratado sobre las costumbres populares de la Mancha. Imaginemos la desesperación del bibliotecario cuando tenga que clasificarlo.
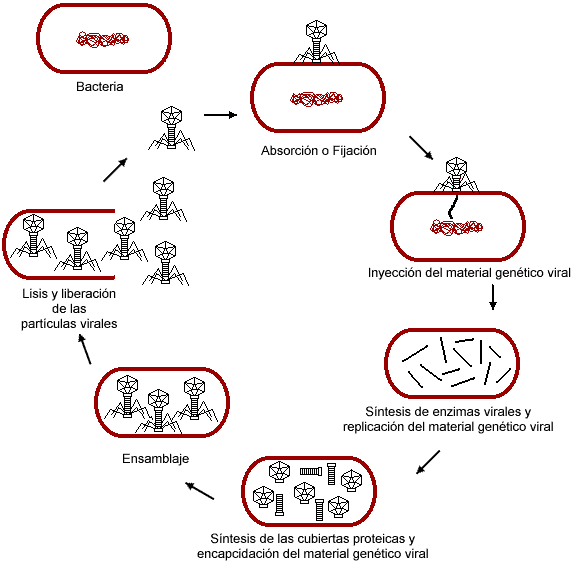
El caso es que algo similar está ocurriendo con el concepto de "especie" en el campo de la microbiología. Podría decirse que debido a los tiempos de generación tan cortos de los microorganismos se están dando cambios evolutivos en tiempo real. En la entrada dedicada a los experimentos de Richard Lenski ya vimos que se había detectado el cambio evolutivo en una población bacteriana estudiada continuamente durante 19 años. Si hacemos una pequeña extrapolación e imaginamos que el experimento de Lenski hubiese comenzado en 1885, año en el que fue aislada y descrita la bacteria Escherichia coli, la cepa evolucionada habría sufrido al menos 6 cambios importantes con respecto a la cepa parental. Pueden parecer pocos, pero ahora consideremos esto:
Este es el perfil de reacciones diagnósticas para diferenciar al género Escherichia del género Enterobacter.

Si nos fijamos, la diferencia entre ambos géneros se reduce a las seis pruebas señaladas en negrita. Es evidente que las diferencias entre esos dos géneros son mayores que tan sólo estos seis cambios, pero también es cierto que cualquier microbiólogo consideraría que estaría ante dos especies distintas si encontrara esas seis diferencias entre dos cepas de bacterias.
Pero lo que ha acabado de dar la puntilla al concepto de especie ha sido la aplicación de las técnicas moleculares en la Ecología Microbiana. Tema que será desarrollado en el siguiente comentario del blog.






































{kind=link}
{kind=link}